Assim como o êxito académico, a vitória numa maratona importante só vem após trabalho árduo. Os músculos das pernas de maratonistas de elite contêm mais mitocôndrias do que os da maioria das pessoas.
A energia química libertada pela hidrólise de ATP nessas mitocôndrias pode ser convertida em energia mecânica para movimentar os músculos.
As células do tecido muscular reúnem-se em dois tipos de fibras.
A maioria das pessoas possui proporções aproximadamente iguais de fibras de cada tipo. Todavia, nos maratonistas de topo, 90% dos músculos do corpo possuem as assim chamadas fibras de contração lenta. As células dessas fibras têm muitas mitocôndrias e utilizam oxigénio para decompor gorduras e glícidos, formando ATP.
Por outro lado, os músculos de velocistas são formados por aproximadamente 80% de fibras de contração rápida, que apresentam menos mitocôndrias. As fibras de contração rápida geram "explosões" de ATP na ausência de O2, mas o ATP é logo utilizado. Pesquisas extensivas com atletas mostram que o treino pode melhorar a eficiência da circulação do sangue nas fibras musculares, proporcionando mais oxigénio e até mesmo uma mudança
da relação entre fibras de contração rápida e fibras de contração lenta.
Agora entra o rato da maratona. Não se trata de um desenho animado ou jogo de computador, mas um rato bem real, programado geneticamente por Ron Evans, do Salk Instituto, para expressar níveis altos da proteína PPARd nos seus músculos.
Essa proteína normalmente controla a decomposição de gordura em tecidos adiposos e também está presente em músculos de contração lenta, onde estimula a decomposição controlada de gorduras para produzir ATP. O rato de Evan foi concebido para decompor melhor as gorduras e, portanto, ser mais magro - mas o resultado dessa pesquisa trouxe um bónus inesperado, Com níveis altos de PPARd, houve aumento em fibras de contração lenta e decréscimo em fibras de contração rápida. Era como se, por um longo período, o rato tivesse sido treinado para maratona!
Os ratos maratonistas são mais magros e mais hábeis do que os ratos comuns.
Mais magros, pois são eficientes na queima de gordura; e mais hábeis em termos de capacidade de correr por longas distâncias. Numa roda giratória de exercício, um rato normal pode correr por 90 minutos, aproximadamente meia milha (900 metros), antes de fatigar-se, Os ratos com aumento de PPARd podem percorrer distâncias quase duas vezes mais longas e duas vezes mais rápido - marcas de verdadeiros corredores de longas distâncias. Poderíamos também manipular genes a fim de aumentar o desempenho (e a queima de gordura) em humanos?
A engenharia genética de pessoas, se exequível, provavelmente situa-se num futuro distante. Todavia, o implante de tecido muscular geneticamente alterado de fato não é uma ideia inconcebível, e já tem provocado preocupações quanto ao aumento inadequado do estado atlético, Mais provável, a curto prazo, é o uso de uma droga experimental denominada GW501516 (desenvolvida pela indústria Glaxo-Smith/Kline), que ativa a proteína PPARd, Ao utilizar a droga em ratos normais, Evans e colaboradores alcançaram os mesmos resultados obtidos com ratos geneticamente modificados. Por estimular a decomposição de gorduras, essa droga está a ser testada no tratamento da obesidade. A energia armazenada no ATP é a que você sempre utiliza para suprir ações conscientes, correr uma maratona ou clicar em links deste post, por exemplo, e ações automáticas do seu corpo, como respirar ou contrair os músculos do coração.
Texto Adaptado de : Vida, a ciência da Biologia. Artmed




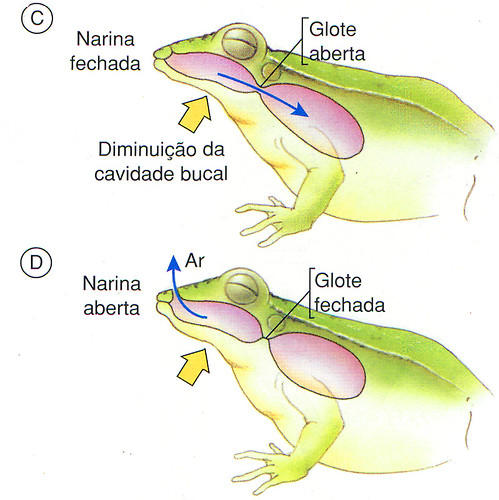 Graças à presença da glote, uma válvula localizada na laringe, o ar pode ser mantido nos pulmões dos anfíbios por um certo tempo. Enquanto isso, eles abrem as narinas, eliminando o ar da boca e captando ar novo, para recomeçar outro ciclo de ventilação pulmonar.
Graças à presença da glote, uma válvula localizada na laringe, o ar pode ser mantido nos pulmões dos anfíbios por um certo tempo. Enquanto isso, eles abrem as narinas, eliminando o ar da boca e captando ar novo, para recomeçar outro ciclo de ventilação pulmonar.











